My first research post will be on a study we recently published. You can access that paper here or here. This project is really a culmination of a lot of my previous work, so it’s a bit backwards to be featuring it first. Where it builds of off past studies I’ll highlight those, and I may do a couple throwback Thursday posts along the way, too. Although I try to write for a general audience, there’s no avoiding some of the jargon that makes its way in. This handy guide will help you decipher some of the technical blabber.

The project really owes its conception to Federico Fanti (above on the left) and Phil Bell. In 2016, I joined them in the field in the Nemegt Basin in the Gobi Desert of Mongolia, and loose threads about a special issue on the Nemegt locality began to tighten. As a bit of background, the Nemegt Basin is a series of eleven Cretaceous localities (~70-66 million years old) exposed in the Gobi Desert of Mongolia. These sites were first discovered by Russian expeditions, and followed up by Polish expeditions. Federico and Phil envisioned a series of papers honouring work done in the past, and advancing our knowledge of these fantastic fossil sites using new methods. I was honoured to be invited to contribute a paper on the oviraptorosaurs of Nemegt. Unfortunately, we had recently published an article on a bonebed of Avimimus from the area, which would have been a perfect piece for the issue. But, in looking at the material for that study, I noticed some anatomical discrepancies that made me suspicious that not all Avimimus material was really the same species. This started a chain reaction that, over the next 8 months, would snowball into a gargantuan task.
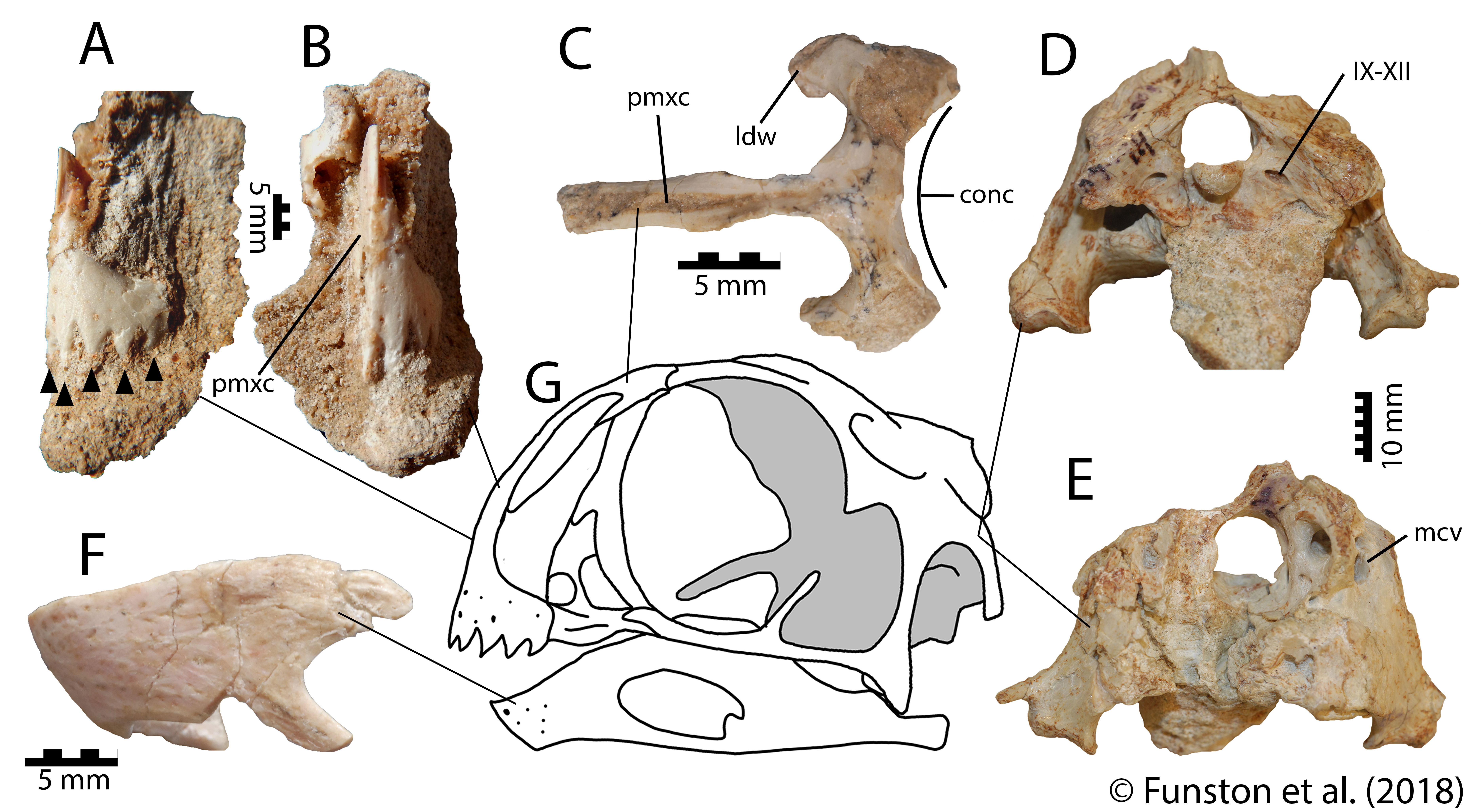
I had already begun several small projects redescribing the holotypes of several Mongolian oviraptorids. Among these were Conchoraptor, Heyuannia (=’Ajancingenia‘=’Ingenia‘), and Rinchenia. These three taxa, despite being some of the first oviraptorids described, are among the most poorly known. Because these three were from the Nemegt Basin, I decided that I would use this opportunity to revise, redescribe, and update our knowledge of their anatomy. This would put new information out there that could be used to improve our phylogenies. The manuscript was starting to take shape: first, an anatomical section naming the new species of Avimimus and reviewing the oviraptorid taxa from the Nemegt Basin. The second section was where I could expand and add some ecological insights that would make the paper appropriate for the special issue.
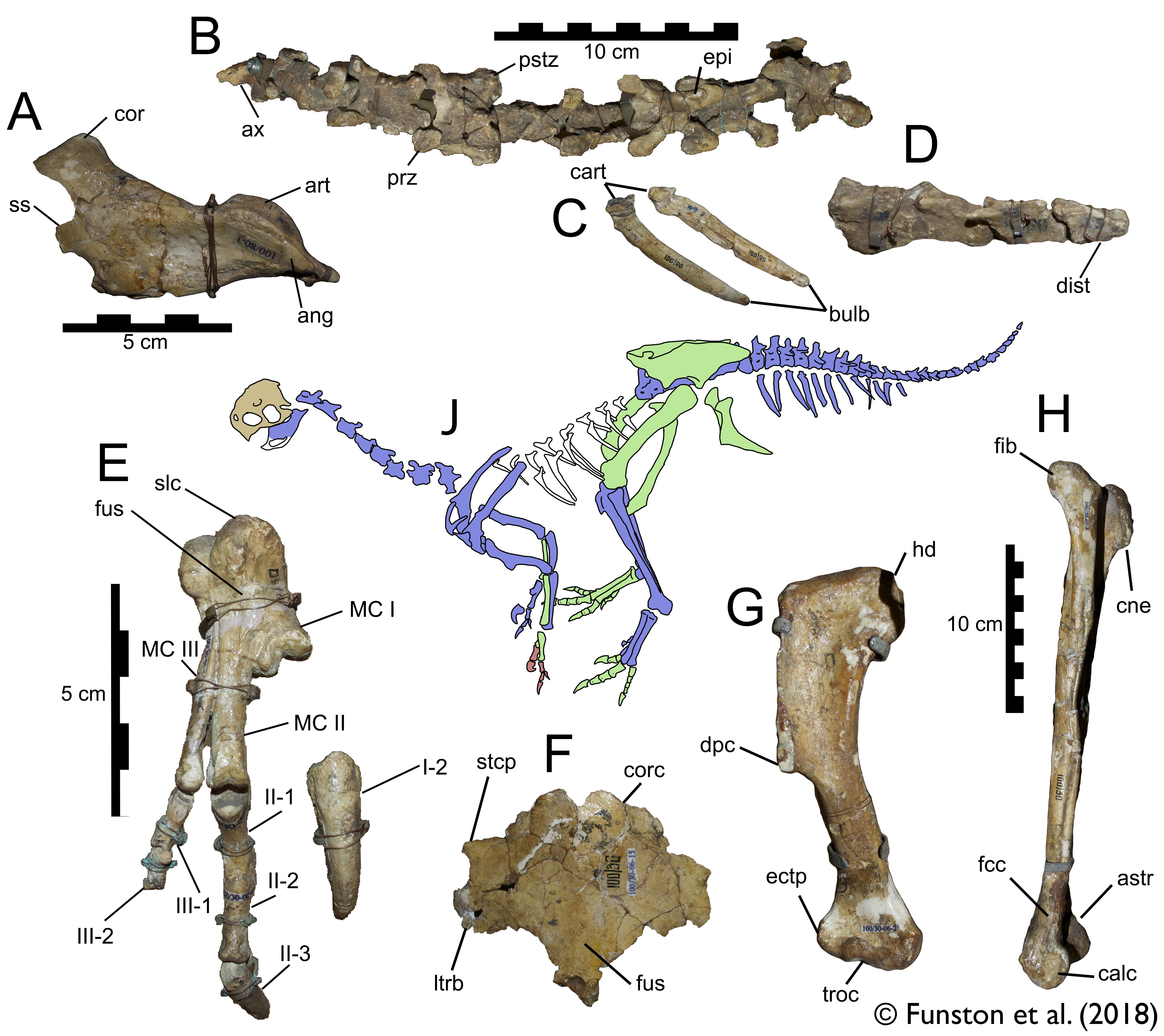
The anatomical section was completed fairly quickly, because it was mostly data I had already collected. So, I turned to a couple questions that had slowly been pestering me more and more. Why were oviraptorosaurs so diverse in this part of the planet, and how could they all coexist? There were at least 5 different genera of oviraptorids in the Nemegt region, and they coexisted with at least one caenagnathid (maybe more), and an avimimid. And that’s ignoring their overlap with other toothless or beaked theropods like ornithomimids and therizinosaurs. Another one of the big questions I had was about the distribution of oviraptorosaur fossils. Most species appear only at one locality, and are represented by multiple specimens there. For example, Nemegtomaia is only known from the Nemegt locality, but there are 3 specimens there. Why aren’t these three specimens spread out between localities? Definitive Conchoraptor material is only known from Hermiin Tsav, and as many as 8 skeletons have been found there. A new oviraptorid seems to be known from Bugiin Tsav and Guriliin Tsav, which are neighbouring localities, based on 6 skeletons. Is this a true pattern, and if so, what causes it?
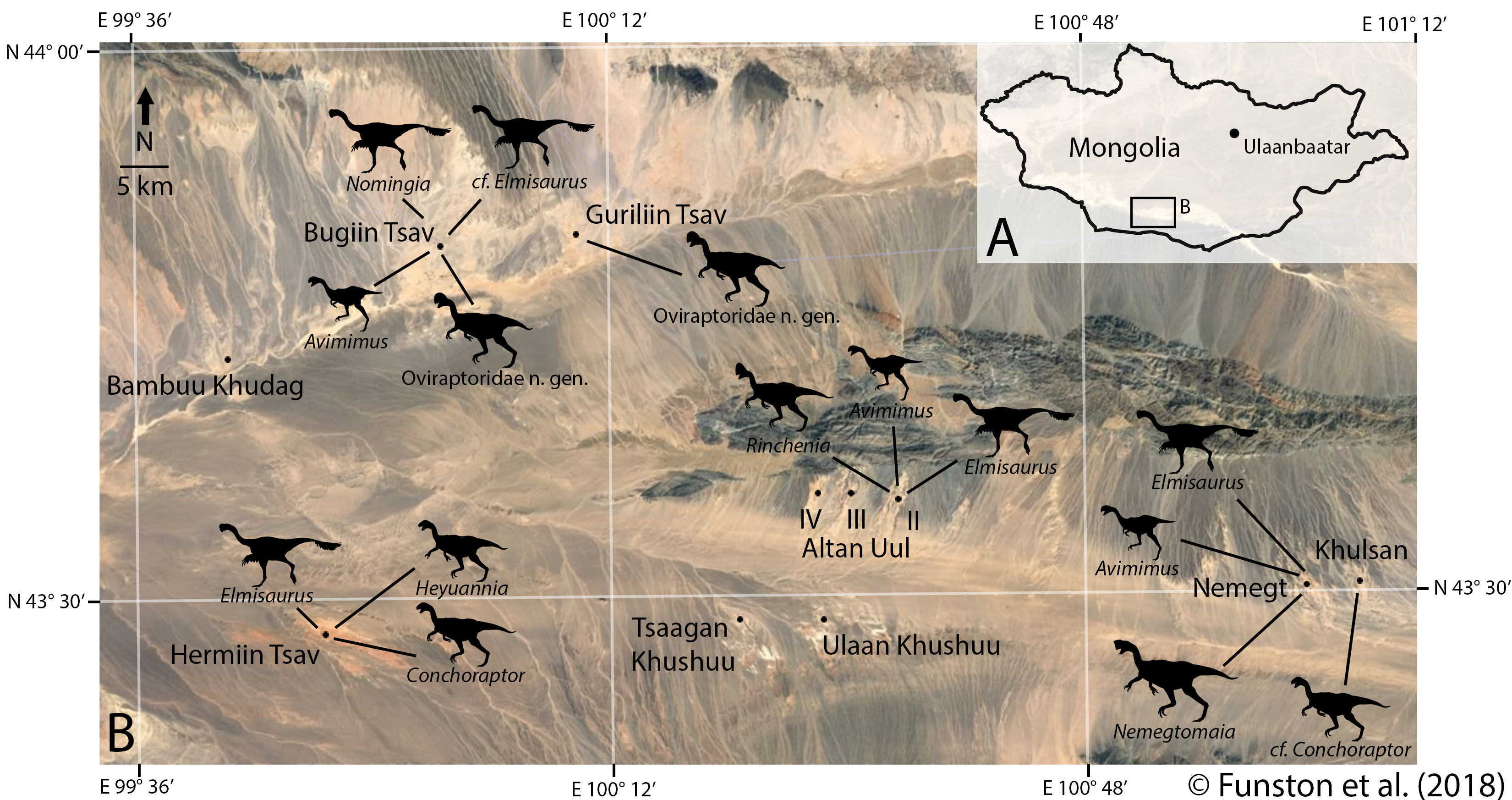
These weren’t questions that could be easily answered using anatomy, so I turned to my friend and colleague Steven Mendonça, with whom I’d had some conversations about testing vertebrate community ecology. Together, we compiled and analyzed a dataset of nearly 500 skeletons that would allow us to test some ecological hypotheses. We treated each locality as its own community, and counted how many of each kind of dinosaur had been found there. Most of our analytical methods would come from invertebrate palaeontology, which is blessed by large sample sizes. Although 500 skeletons is nothing to scoff at, these skeletons are unevenly distributed between 11 localities. So, we first had to test how good our samples were. That way, we could understand whether we could trust any signal we found.
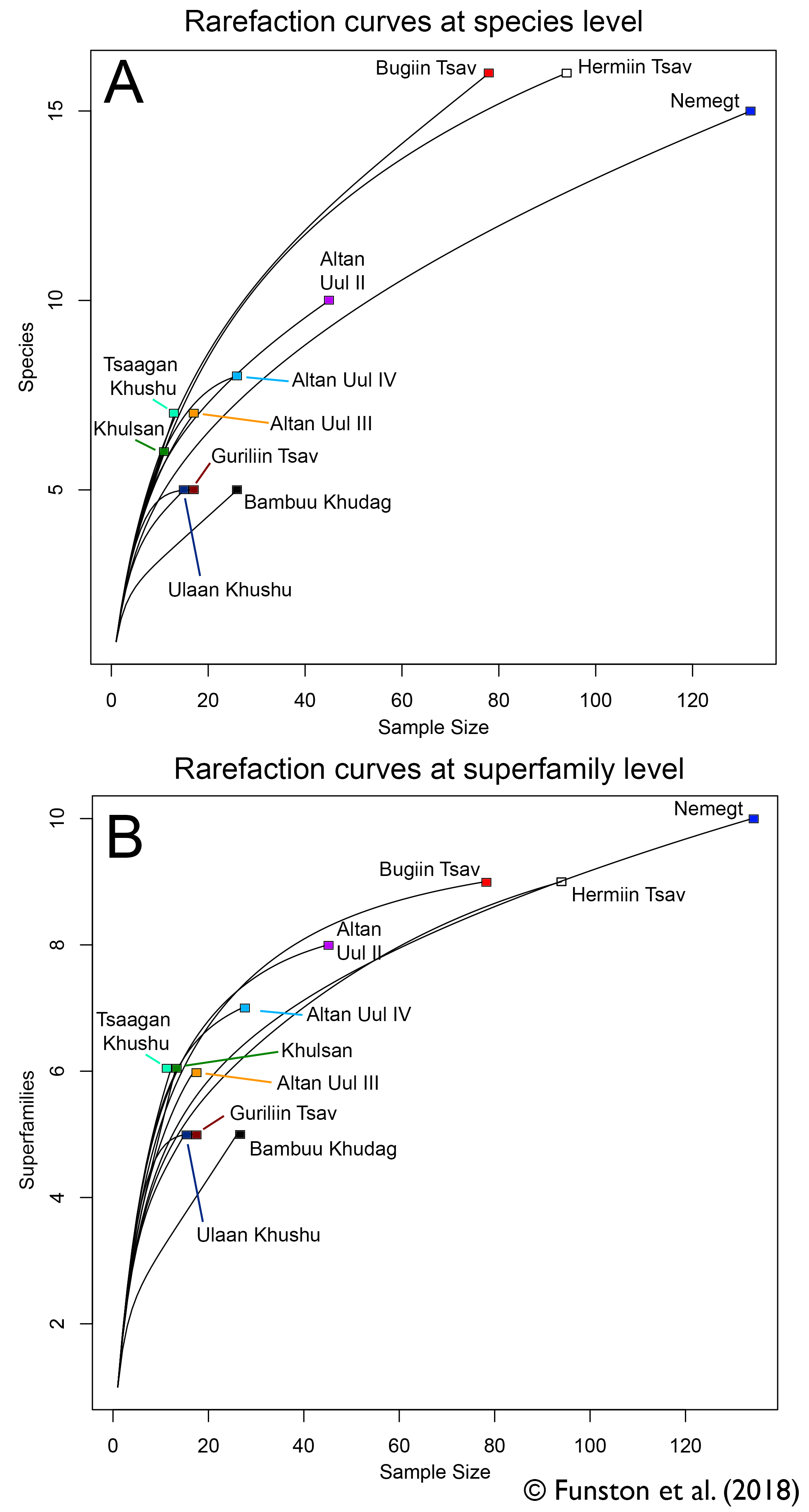
We used a method called rarefaction, which randomly and iteratively subsamples our data. This determines at what point we reliably stop finding new taxa, and if we’ve reached that point. The results showed that while most of the sites were still very poorly sampled, we might be able to get a signal from some of the more intensively worked areas. These were Altan Uul II, Bugiin Tsav, Hermiin Tsav, and Nemegt. Luckily, these are the sites that have produced nearly all the holotype material for Nemegt oviraptorids. They are also the sites that are best constrained geologically, which is promising. So, the pattern of oviraptorid distribution might be a real one.
Our other analyses showed that communities throughout the Nemegt interval were fairly stable. That’s to say, if there was change over time or space, it didn’t affect the overall community structure. So, these oviraptorids aren’t members of different communities. The next step is where Steven’s invertebrate ecology wizardry came into play. He developed a test that pitted our data against randomly generated datasets derived from our data. This allowed us to test whether our fossil finds were evenly distributed. Most dinosaurs were evenly distributed: the number of specimens is a function of the sample size. This was true of most oviraptorosaurs too, except for the oviraptorid species. This gave us some support that the oviraptorid species were actually clumping together. We settled on two possible explanations: the oviraptorid species were separated either spatially or temporally. Unfortunately, this is as far as the data could take us. The geology of the Nemegt interval is still poorly understood, and we can’t be certain how much time the rocks represent. As of yet, there’s no way to test whether the trend we’re seeing is spatial or temporal.
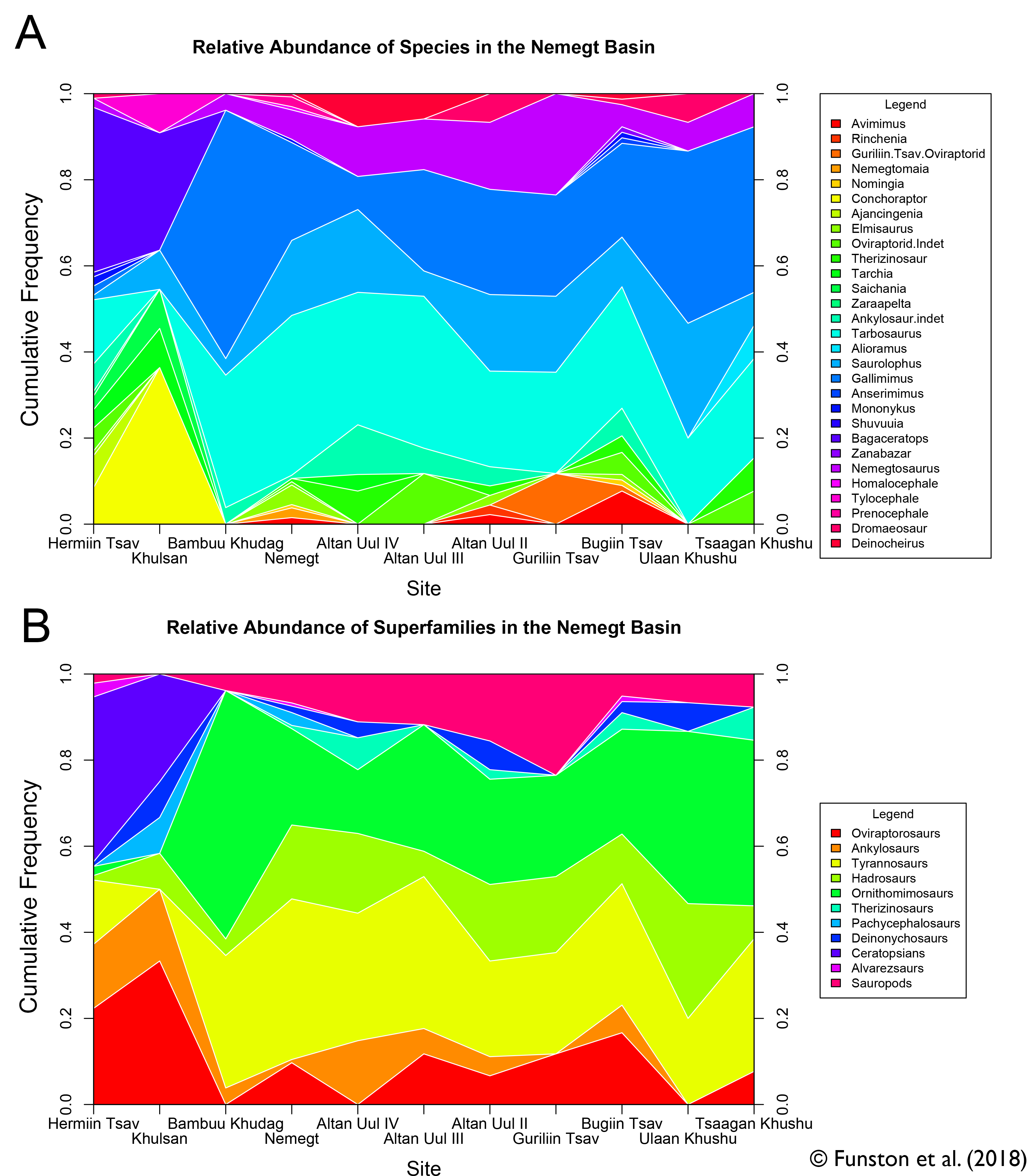
This led us to conclude that oviraptorids probably coexisted by not coexisting: the diversity was the result of either spatial segregation, or rapid species turnover. The data helped us understand how avimimids, caenagnathids, and oviraptorids all coexisted, too. First, there’s a clear difference in habitat preference between avimimids, caenagnathids, and oviraptorids. The first two groups are exclusively found in the rocks deposited by rivers and streams. Oviraptorids are more common in rocks that represent ancient dunes. However, oviraptorids seem to be less sensitive to habitat change, because they are found in both types of rocks. Our data also shed light on the community roles oviraptorosaurs may have played. As herbivores there was lots of competition for food, and their small numbers probably meant that they were specialized for a certain type of food. Similarities to modern parrots make me think this may have been fruits and hard nuts.
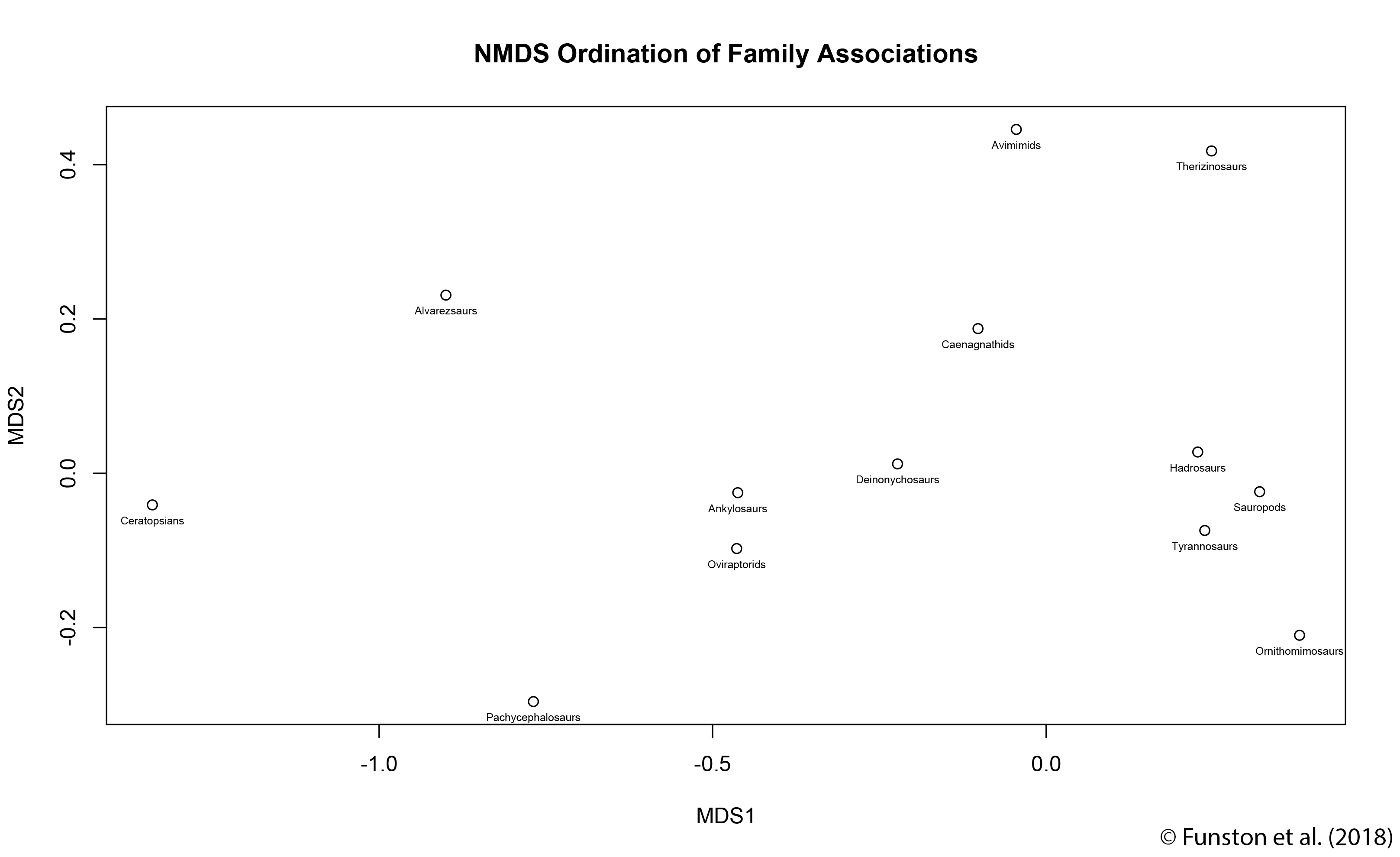
The results of the paper help us understand the anatomy and ecology of oviraptorosaurs, and hopefully provide a good foundation for future work. This paper is a tome of information for the interested reader, and unfortunately I can’t cover all of our findings here. If I had to pick one advance that I think is important, it would be the creation and analysis of this kind of dataset. Typically vertebrate datasets are “n < 1”, where you’re working off of incomplete skeletons, or are generated from microsites where most taxa can’t be identified to a species level. Having access to species-level data with such a large sample size is really powerful, as is being able to apply statistical tests typically reserved for invertebrate palaeontology. This dataset was only possible because of decades of hard work by Phil Currie, who collected the vast majority of the data using a handheld GPS. His foresight in collecting data even when its use wasn’t immediately obvious is what made our project possible. An important lesson for us all!
-Greg

3 Replies to “Oviraptorosaurs of the Nemegt”